Conceito de Músculo
Os
músculos formam o maior conjunto de tecidos do organismo e representam
cerca de metade do peso do corpo. O tecido muscular tem origem
mesodérmica e é composto por células alongadas, denominadas células
musculares, miócitos ou fibras musculares, arranjadas em feixes
revestidos por tecido conjuntivo e especializadas na contração. No
músculo, a membrana celular chama-se sarcolema e o citoplasma é o
sarcoplasma.
Existem
três tipos de tecidos de musculares classificados de acordo com as suas
características morfológicas e funcionais (Fig. 1). Distinguem-se os
músculos estriados esqueléticos e cardíaco, que apresentam uma estriação
transversal quando observados ao microscópio ótico, e os músculos lisos
(sem estriação). Por outro lado, fala-se em músculos voluntários
(esqueléticos) e involuntários (lisos e cardíaco) consoante estejam
enervados respetivamente pelo sistema nervoso somático ou pelo sistema
nervoso autónomo.
Algumas
propriedades são partilhadas pelos três tipos de tecidos musculares: O
sistema contrátil é constituído por filamentos finos que deslizam entre
filamentos espessos e a fonte de energia utilizada pelos três é o
trifosfato de adenosina (ATP). Contudo, a estrutura e a organização das
fibras e o modo de excitação são diferentes nos três casos.
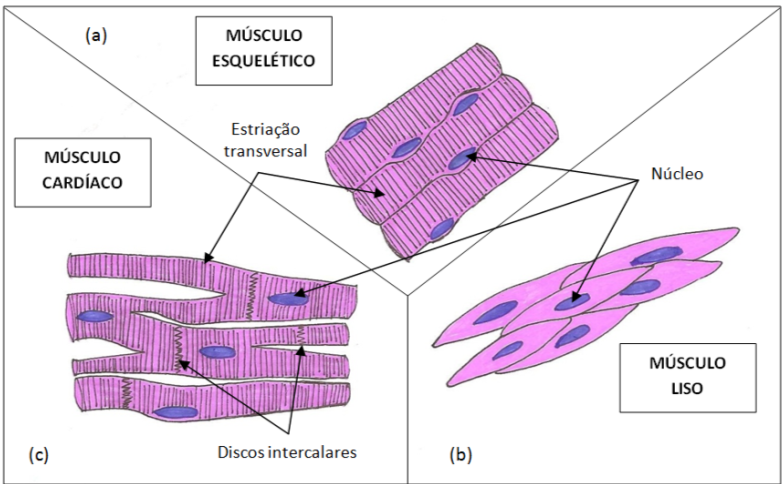
Fig.1.
Representação dos diferentes tipos de músculos (Ilustração de Anabela
Fernandes, 2015)
Músculo esquelético
As
células do músculo esquelético são longas e cilíndricas. Apresentam
vários núcleos localizados na periferia próximos da membrana celular
(Fig. 1a). O sarcolema é envolto por uma lâmina basal e células
satélites. Este possui invaginações profundas, designadas por túbulos
transversos ou túbulos T, envolvidas na rápida despolarização da
membrana. Cada fibra, caraterizada por possuir múltiplas miofibrilas,
está envolta por uma delicada camada de fibras reticulosas, designada
endomísio, na qual existe uma extensa rede de capilares sanguíneos e de
nervos. As fibras musculares juntam-se em feixes ou fascículos envoltos
por uma bainha fibro-elástica, o perimísio. Um conjunto de feixes
musculares forma assim um músculo rodeado, por sua vez, por uma camada
densa de tecido conjuntivo designada epimísio.
A
maioria
dos músculos esqueléticos, no ser humano, é formada por uma combinação
de diferentes tipos de fibras musculares. As três principais categorias
de fibras são:
- As
fibras
do tipo I: Fibras vermelhas aeróbias de contração lenta e resistentes à
fadiga cujo processo metabólico preferencial é a fosforilação oxidativa;
- As
fibras
do tipo IIa: Fibras intermédias, fibras de contração rápida e
moderadamente resistentes à fadiga, com alta atividade glicolítica e
oxidativa;
- As
fibras
do tipo IIb: Fibras brancas anaeróbias de contração rápida, bastante
sensíveis à fadiga, que usam preferencialmente a via glicolítica.
A
miofibrila,
elemento contrátil especializado da fibra muscular, é composta por
elementos do citoesqueleto dispostos de forma regular: os filamentos
espessos e finos. A unidade funcional deste sistema é o sarcómero (Fig.
2).
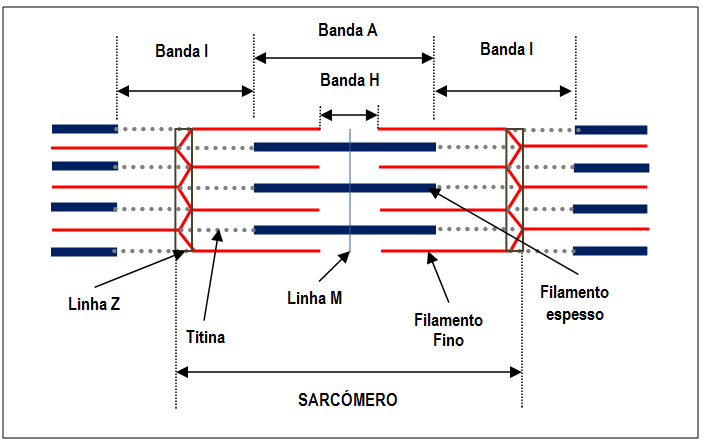
Fig. 2.
Representação esquemática de um sarcómero no estado relaxado (Ilustração
de Anabela Fernandes, 2015)
As
miofibrilas
apresentam uma alternância de bandas escuras (banda A) e bandas claras
(banda I). Estas bandas encontram-se alinhadas dando um aspeto estriado
ao músculo esquelético. As bandas A, tal como representadas na figura 2,
são constituídas por filamentos espessos inteiros e sobreposições de
parte dos filamentos finos. Por sua vez, as bandas I são constituídas
unicamente de filamentos finos.
Os filamentos espessos
são compostos por várias centenas de proteínas de miosina e os
filamentos finos são constituídos por três proteínas, a actina
(constituinte mais abundante), a troponina e a tropomiosina. Cada
molécula de actina possui um local
de ligação para
as pontes cruzadas da miosina.
A elasticidade do
músculo e a estabilidade dos filamentos finos e espessos são garantidas
através da presença das proteínas da linha
M e de uma
proteína elástica gigante, a titina. Esta proteína, a maior do organismo
(cerca de 30.000 aminoácidos), encontra-se ligada aos filamentos de
miosina e estende-se da linha Z à linha M.
Mecanismo de
contração muscular
A estimulação do
músculo esquelético é devida à libertação de um neurotransmissor, a
acetilcolina, na junção neuromuscular (junção ente a parte terminal de
um neurónio motor e a placa motora da fibra muscular). A ligação da
acetilcolina aos recetores na placa motora leva à propagação de um
potencial de ação (PA) na superfície do sarcolema e penetra no interior
da
fibra através dos túbulos T.
O retículo
sarcoplasmático forma uma rede descontínua de túbulos finos que envolvem
as miofibrilas. As extremidades de cada segmento são dilatadas, formando
cisternas laterais que funcionam como reservatórios de cálcio (Ca2+).
Quando o PA
chega aos túbulos T,
este desencadeia a abertura dos canais de Ca2+. O Ca2+
libertado no citosol liga-se à troponina provocando o deslocamento da
tropomiosina, afastando-a do local de ligação das pontes cruzadas da
miosina na actina. As pontes cruzadas executam então um movimento de
força que permite o deslizamento dos filamentos de actina e miosina.
Vários ciclos de contração acontecem enquanto os locais de ligação
estiverem expostos e o Ca2+ disponível.
Quando os
níveis
de cálcio citosólico diminuem, o Ca2+ desliga-se da troponina
e a tropomiosina volta à posição original resultando no relaxamento do
músculo.
A energia mecânica
necessária
ao processo de contração, nos três tipos de tecidos musculares, resulta
da transformação da energia proveniente da hidrólise do ATP. No músculo
esquelético, esta hidrólise requer a ligação do magnésio (Mg2+)
ao ATP pois o Mg2+ funciona como acelerador da atividade da
ATPase da miosina. Esta enzima cliva o ATP em difosfato de adenosina
(ADP) e fosfato inorgânico (Pi) (reação de hidrólise do ATP). A energia
química libertada é armazenada nas pontes cruzadas e transformada em
energia mecânica necessária à ligação da miosina na actina e ao
deslizamento dos miofilamentos. O ADP e o Pi são rapidamente libertados
pela miosina. A ATPase torna-se então disponível para a ligação de uma
nova molécula de ATP, o que leva à rutura da ligação entre a actina e a
miosina, voltando ao início do processo.
Quando
a atividade muscular é intensa, a quantidade de ATP, proveniente da
respiração celular e presente na célula muscular, não é suficiente. A
principal fonte de reserva energética do músculo é então o fosfato de
creatina. A creatina quinase transfere um grupo fosfato do fosfato de
creatina para o ADP formando ATP e creatina. Por outro lado, quando há
um excesso de ATP, a reação inversa é catalisada, transferindo um grupo
fosfato do ATP para a creatina, formando de novo o fosfato de creatina.
Consequentemente, a quantidade de ATP livre no músculo durante uma
contração não se altera muito.
Curiosidades:
- No homem, as
fibras musculares são mais espessas e os músculos mais desenvolvidos do
que na mulher. Isto deve-se à testosterona, uma hormona esteroide
secretada em muito maior quantidade no homem e que favorece a síntese da
actina e da miosina.
- Depois da morte,
a ausência de ATP disponível faz com que as ligações actina-miosina
persistam, impossibilitando os ciclos de contração-relaxamento. Este
fenómeno produz a rigidez cadavérica ou rigor mortis. A rigidez
inicia-se entre 3 e 4 horas após a morte e é completa após 12 horas. O
relaxamento dá-se após aproximadamente 36 horas.
Músculo liso
O
tecido
muscular liso encontra-se em camadas ou feixes nas paredes do intestino,
do ducto biliar, da bexiga urinária, do trato respiratório, do útero,
dos vasos sanguíneos, na íris e corpo ciliar, no músculo eretor do pelo,
entre outros. As células deste tecido são mais curtas do que as células
do músculo estriado, são fusiformes e possuem um núcleo central (Fig.
1b).
As
células
do tecido muscular liso possuem três tipos de filamentos: os filamentos
espessos de miosina, mais compridos do que no músculo esquelético, os
filamentos finos de actina e tropomiosina mas desprovidos de troponina e
os filamentos intermédios, compostos por desmina e vimentina. No caso do
músculo liso, estes três tipos de filamentos não se organizam em
miofibrilas nem em sarcómeros. Como tal, o músculo liso não apresenta
estriações.
Os
filamentos
intermediários inserem-se no citoplasma e na membrana plasmática através
de estruturas denominados corpos densos. Por outro lado, a membrana
possui uma grande quantidade de depressões de pequenas dimensões, as
cavéolas, que atuam como um sistema primitivo de túbulos T e são
responsáveis pelo transporte, por pinocitose, dos iões Ca2+.
A
miosina
do tecido muscular liso é uma miosina do tipo II, formada por duas
cadeias pesadas e dois pares de cadeias leves. Esta encontra-se
enrodilhada, exceto quando combinada com um radical fosfato que leva ao
desdobramento do filamento.
Após
um determinado estímulo, os níveis de Ca2+ citoplasmático
aumentam. O Ca2+ provém do retículo sarcoplasmático e também
entra a partir do líquido extracelular. A seguir, este
liga-se à calmodulina formando um complexo que ativa a quinase da
miosina. Esta última fosforila a cadeia leve da miosina que se desdobra
expondo o local de ligação da actina na cabeça da miosina. A miosina
pode então ligar-se aos filamentos de actina causando a contração
muscular. Quando os níveis de Ca2+ são reduzidos, a cadeia
leve da miosina é enzimaticamente desfosforilada e o músculo relaxa.
As
células
do músculo liso estão ligadas umas às outras através de junções
comunicantes que permitem a contração sincronizada do músculo liso. Por
outro lado, as células são revestidas por uma lâmina basal e estão
envoltas por uma rede de fibras reticulares que mantem as células unidas
de tal maneira que a contração de apenas algumas células se transforme
na contração do músculo inteiro.
Por
último,
a contração das células do tecidos muscular liso é estimulada mediante
sinais nervosos, por estimulação hormonal, pelo estiramento do músculo,
entre outros meios.
Músculo cardíaco
Este
músculo é específico do coração e é responsável por impulsionar o sangue
através das aurículas e dos ventrículos e para dentro dos vasos
sanguíneos. As células do músculo cardíaco ou cardiomiócitos possuem um
núcleo central, são cilíndricas e ramificadas (Fig. 1c). As células
interconectam-se para formar redes complexas. São células mais pequenas
do que as do músculo estriado, com aproximadamente 15µm de diâmetro e 80
a 100µm de comprimento.
O
músculo
cardíaco possui propriedades de estrutura e função em comum com o
músculo esquelético. As fibras cardíacas estão circundadas por uma
delicada camada de tecido conjuntivo, equivalente ao endomísio do
músculo esquelético, que contém uma abundante rede de capilares
sanguíneos. Tal como no músculo esquelético, o músculo cardíaco é
estriado, todavia as seguintes diferenças são observadas: os túbulos T e
o retículo sarcoplasmático não são tão bem desenvolvidos; os
cardiomiócitos estão unidos pelas extremidades através de conexões
intercelulares especializadas, os discos intercalares, que são
estruturas fortemente coráveis e que apresentam uma organização em
escada, com uma porção transversal, contendo desmossomas, e uma porção
longitudinal onde estão localizadas as junções comunicantes (Fig. 1c).
Esta é uma característica exclusiva do músculo cardíaco.
Contudo,
tal como no músculo liso, a contração do músculo cardíaco é involuntária
e o Ca2+ provém do retículo sarcoplasmático e do líquido
extracelular.
Fontes
Gartner,
L.P. and Hiatt, J.L .
(2012). Histologia Essencial. Rio de Janeiro:
Elsevier Brasil. p132-151.
kierszenbaum,
A.L. and Tres L.L. (2012). Histologia
e Biologia Celular - Uma introdução à patologia.
3rd ed. Rio de Janeiro: Elsevier Brasil. p169-171.
Sherwood,
L. (2006).
Physiologie musculaire. In: De boeck and Larcier.
Physiologie humaine: A Human Perspective. Bruxelles: De boeck.
p203-235.
Widmaier,
E.P. et al.
(2003). Human
Physiology - The mechanisms of body function. 9th ed. Boston:
McGraw-Hill. p376; 450-454.
|
Procure outros termos na nossa enciclopédia
|
A |
B |
C |
D |
E |
F |
G |
H |
I |
J |
K
| L |
M |
N |
O |
P |
Q |
R |
S |
T |
U |
V |
W |
X |
Y |
Z |
|
|



